A trait is any observable characteristic of an organism. Historically, traits were anatomical features, developmental and embryological processes, behavioral characteristics and gene sequences. Prior to the advances in molecular biology, most traits were anatomical / structural features which have the advantage of being preserved in the fossil record. However, the foundation of anatomical features generally is complex and rarely due to a single gene pair as Mendel reported. Over the last fifty years, advances in molecular biology have made gene sequences the traits of choice for studying relationships between living organisms. Of course, rarely will ancient fossil remains contain DNA for comparison with living forms so there is always a combination of traits used in developing phylogenetic trees.
An adaptation is an inherited trait that increases fitness in its current environment. The marvelous match between an organism's traits and the features of its environment are the result of natural selection acting on the palette of traits within the population. To be an adaptation, the trait must be (1) inherited, (2) the result of natural selection, and (3) increase the reproductive success (fitness) of the population.
From classical Mendelian genetics, one trait was always dominant to its paired allele. Gregor was not an honest monk. He never reported results he must have obtained in which the two traits, for example yellow and green peas, appeared to "blend". Always, his reported results demonstrated a simple dominant-recessive relationship between two alleles that controlled a particular trait. The reason that I believe Gregor cheated in reporting his results is that most traits do appear to "blend"; that is, they display incomplete dominance and/or polygenic inheritance in which more that one pair of alleles control the phenotypic outcome. Mendel must have carefully selected the traits he investigated in order to support his hypothesis of how traits were transmitted from one generation to the next.
In the case of crossing true-breeding yellow pea plants and green pea plants, where yellow is dominant to green, all progeny received one yellow gene (Y) and one green gene (y), and expressed a yellow phenotype --not a blend of the two colors. When these hybrid Yy individuals were crossed, three-fourths of the offspring were yellow and one-fourth of the offspring displayed the "hidden" green phenotype not appearing in their parents. Mendel's experiments demonstrated only discrete traits --the characters expressed were either yellow or green seeds, tall or short plants, purple or white blossoms, etc. However, other traits appear to be continuous and vary within ranges between two values. For example, in humans, heights are not discrete but are continuous with heights not limited to 4, 5, or 6 feet, but with graded, intermediate heights above, below and between these values. |
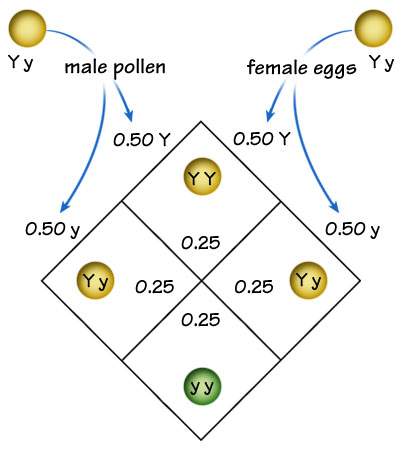
|
When a single pair of alleles govern a trait and one is dominant to the other, simple Mendelian genetics apply and, if the trait is color, there will be only two colors in the population. In Mendel's research this was shown in pea flower color (purple and white) and pea seed color (yellow and green). If the alleles demonstrate incomplete dominance, three colors would be produced. This was not reported by Mendel, but if Mendel's peas had displayed incomplete dominance, pea flower colors would have been seen as purple, violet and white and pea seed colors as yellow, yellow-green and green. These are discrete phenotypes, meaning that there is not a spectrum of colors produced because pairs of individual genes determine the color and the amount of color.
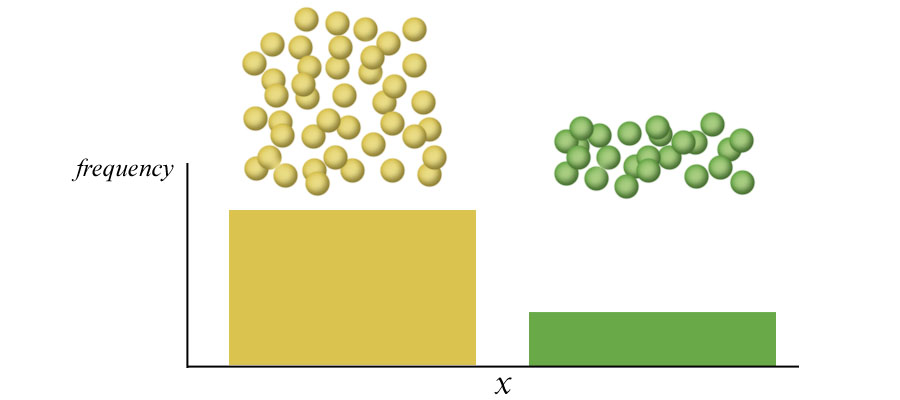
Above. Mendel's peas showed discrete variation with regard to pea color. Peas were either yellow (YY or Yy) or green (yy).
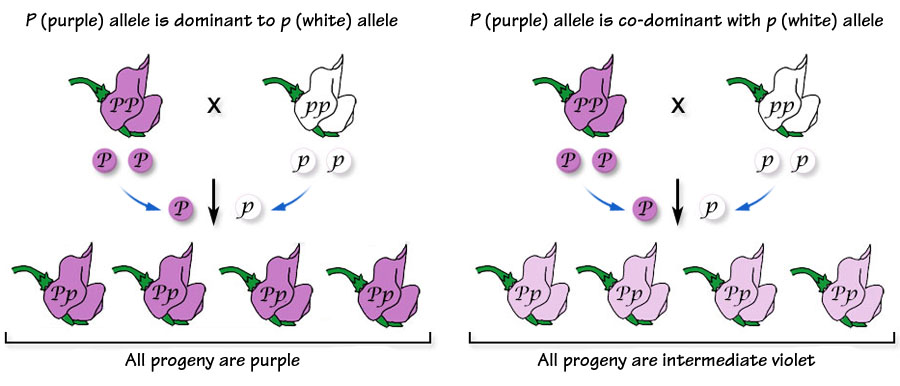
But what if the two alleles were expressed equally? What if the alleles were co-dominant? Recall Mendel's peas that were either purple-flowered or white-flowered. The white-flowered peas contained two alleles for pigment that had been damaged by one or more mutations that prevented the production of pigment. The purple-flowered plants contained one or two alleles that could cause the production of sufficient pigment to produce purple flowers --possessing only one P allele was sufficient. If, however, the purple allele (P) could only cause the production of a limited amount of pigment and that peas required two copies of the allele to produce sufficient pigment to produce a purple flower, then possessing one purple allele and one white allele would be able to produce only half the amount of pigment to produce a purple flower and the flowers would be an intermediate violet color.
Below. Comparing the crosses in which the purple allele (P) is dominant (left) or co-dominant (right). The number of phenotypes increases from two (purple and white) to three (purple, violet and white) when the genes are co-dominant. This affects how the alleles are exposed to natural selection forces. When one allele is dominant it "protects" the recessive allele in the heterozygote; when the two alleles are co-dominant, both alleles are exposed in the heterozygote.
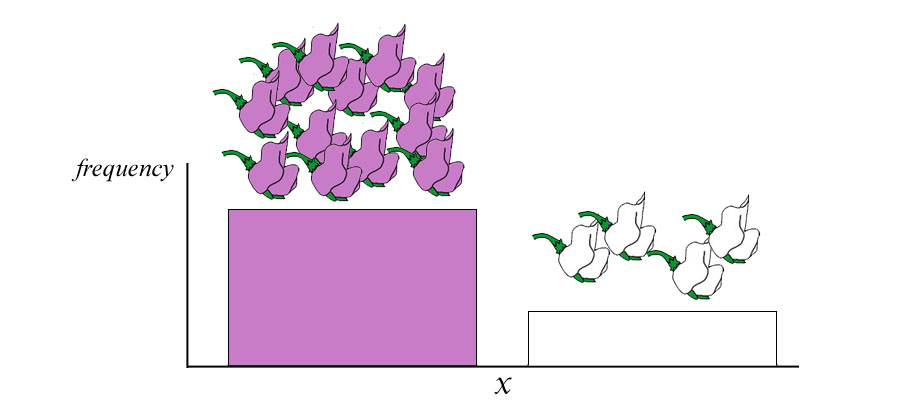
| Below left. If the purple allele is dominant to the white allele, then crossing a true-breeding purple- (PP) with a true-breeding white- (pp) flowering plant will result in progeny all of which are purple despite having both a purple and white allele. | | Below right. If the purple allele is co-dominant to the white allele, then crossing a true-breeding purple- (PP) with a true-breeding white- (pp) flowering plant will result in progeny all of which are an intermediate violet-flowered plant. |
Above. Regardless of whether P is dominant or codominant, the results of crossing a true-breeding purple plant with a true-breeding white plant produces only a single phenotype. In both scenarios, the white phenotype disappears and is protected from forces of natural selection. In the scenario in which P is co-dominant with p, both parental types are hidden and only an intermediate form is expressed. Because the alleles of P and p are discrete "factoren" (genes) and do not "blend", the parental phenotypes can be expressed in future generations --even in the situation where it appears the phenotypes of purple and white have blended to form violet flowers.
Above. In the case of a dominant-recessive allelic system, having the phenotype white (above left) also discloses the genotype, pp, (above right). However, viewing a purple flower one cannot tell whether it is purple due to the genotype PP or the genotype Pp without completing a test-cross, for example a cross with a pp individual and viewing the progeny. In such a test cross, if the purple-flowered plant were homozygous for P, then all the test-cross progeny also would be purple with the genotype Pp. If, on the other hand, the purple-flowered plant were a heterozygous, Pp, then half of the progeny would be purple-flowered (Pp) and half white-flowered (pp).
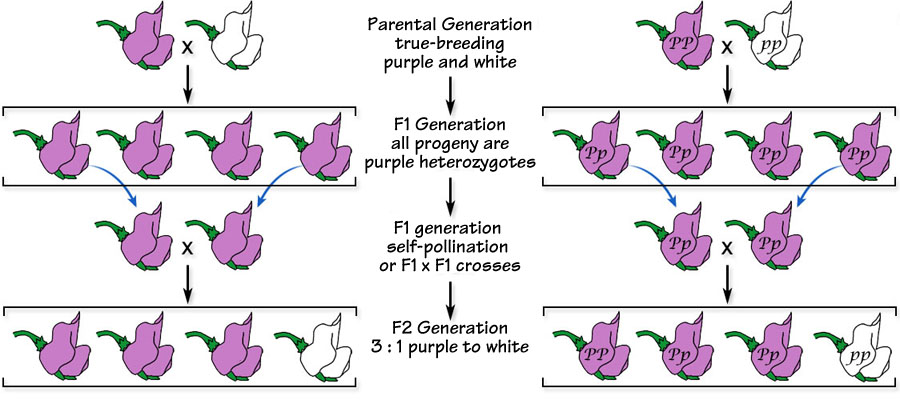
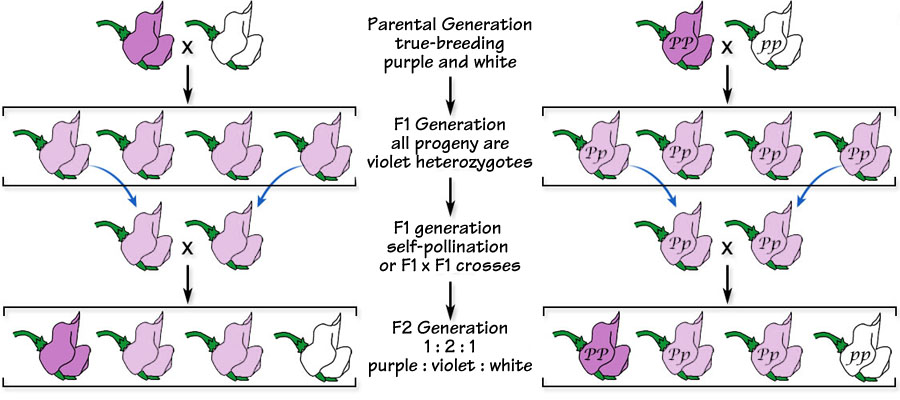
In the F2 generation produced by crossing two purple hybrid flowers (below left), white re-appears; and, in the F2 generation produced by crossing violet plants demonstrating co-dominance (below right), both purple and white re-appear.
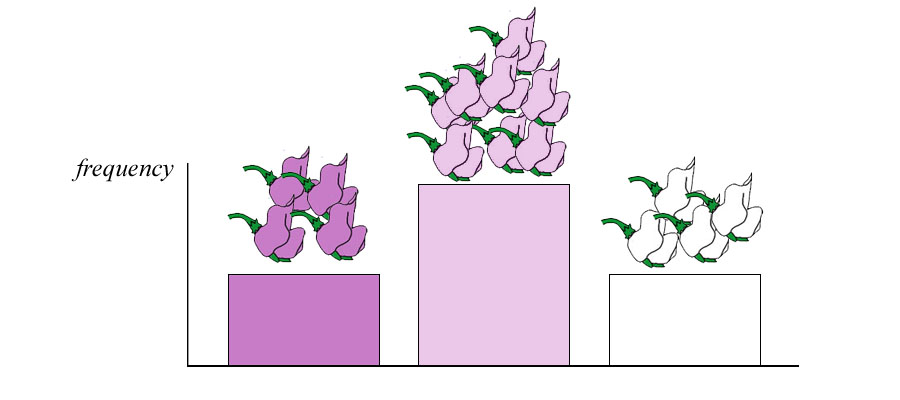
Below. Continuing the co-dominant hybrid crosses by first crossing true-breeding purple- and white-flowered plants and then allowing the hybrids to cross results in an F2 generation that contains purple-, violet- and white-flowered plants in the ratio of 1:2:1. With co-dominance, three phenotypes exist in the population.
Above. In this cross, only one allele pair is responsible for the flower-color phenotype. Crosses in which only one allele pair is responsible for a particular phenotype is referred to as a monohybrid cross. Mendel never reported phenotypes that were determined by more than one allele pair. He investigated plant form (tall vs short), flower position (axial vs terminal), seed color (yellow vs green), seed form,(smooth vs wrinkled), flower color (purple vs white), pea pod form (inflated vs constricted), and pod color (green vs yellow). And, all of the allele pairs he reported exhibited a dominant-recessive relationship.
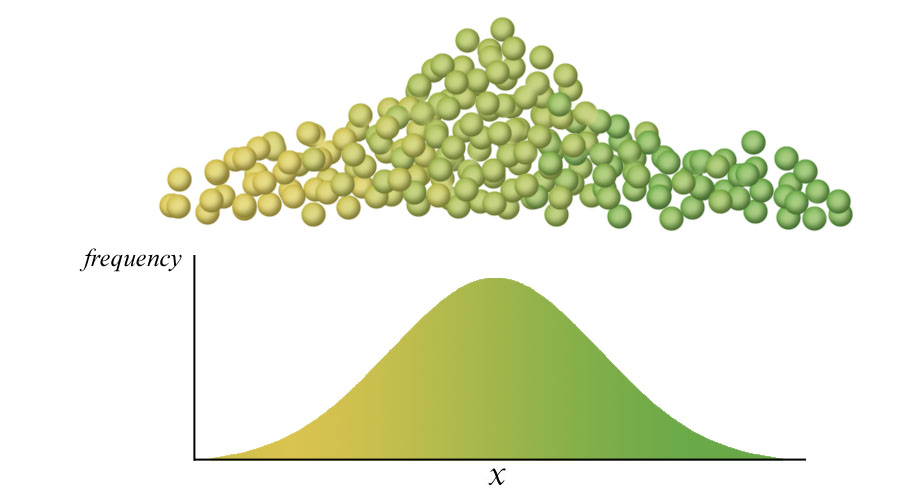
Below. A more usual condition is that a phenotype shows co-dominance, is under control of more than two alleles for any one gene, and often is under control of more than one gene (polygenic inheritance). In addition, quite often the presence of one gene will modify the activity of other genes (pleiotropy), making phenotypes extremely complex and appearing to have continuous gradations rather than discrete characteristics. If polygenic inheritance controlled seed color in peas, then the distribution of seed color in the population would look something like the illustration below.
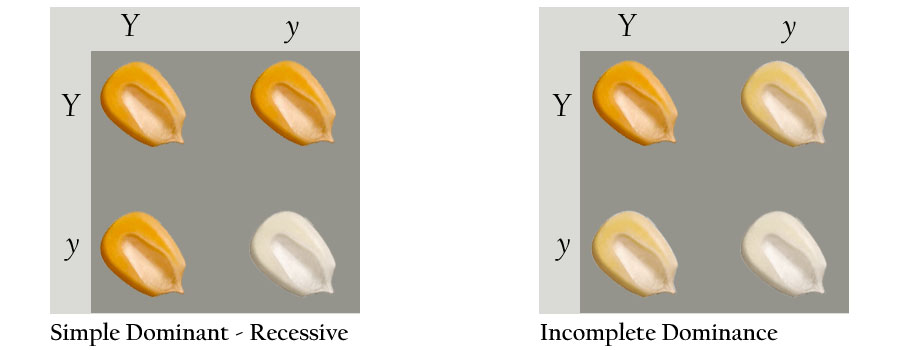
It doesn't take many genes to produce a condition that appears to be continuous instead of discrete. Let us assume that corn seeds are controlled by two alleles, yellow (Y) and white (y). If the two alleles exhibited a simple dominant-recessive condition, there would be two colors of corn seeds (below left). If the two alleles were co-dominant, then three colors of corn seeds would appear in the population (below right).
On the other hand, if seed color were controlled by two genes, each with two alleles, then the variety of seed colors would appear to approach a continuous distribution. In the illustration below, yellow (Y) is co-dominant with white (y), but a second gene for carotenoid pigment also influences color. This gene also has two alleles that are co-dominant, carotenoid orange (C) and white (c). The result still is a discrete distribution of different colors, but the number has increased such that the differences between colors appear to result in a continuous distribution.

The more genes and the more alleles involved in producing a phenotype increases the apparent continuous distribution and, of course, there are environmental influences on phenotypes. Human skin color is an excellent example. Color-producing cells of skin contain pheomelanin and eumelanin, each of which is under the control of multiple co-dominant alleles. Both the amount of each pigment and the color-produced by one or another of the many alleles determines the color of the skin. And, production of pigment can be influenced by other genes that respond to the amount of UV radiation absorbed by the skin --tanning.

|
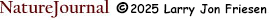 |  |
|