|
|
| Heterospory, Steles and Leaves: Adaptations for Life on Land |
|
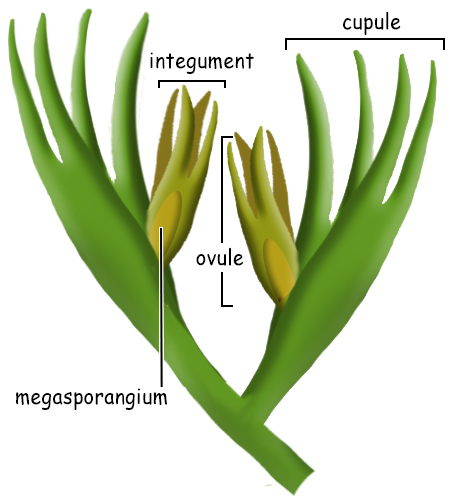
The evolution of the seed is the single most important adaptation of the embryophytes for their transition to a fully terrestrial plant life history. The early steps in this transition that allowed the production of the seed began early in the Devonian with the change from homospory to heterospory. But other adaptations complemented the evolution from homospory to heterospory that allowed not simply the transition to land, but to towering seed plants in vast forests. Homospory to Heterospory The transition from small spores of uniform size to two sizes of spores, large megaspores and small microspores, indicated more than simply differences in spore size. Heterospory indicates a major genetic transformation that changed how sex was determined in plants. The homosporic condition leads to spores that are all the same size, but also the same genetic composition. Spores from homosporous plants germinate to form gametophytes that are potentially bisexual and that can produce both female archegonia and male antheridia. Many homosporous plants have gametophytes that, in fact, are bisexual and do produce both archegonia and antheridia. In others, although the potentual for the bisexual condition exists, the environment influences whether the gametophyte will produce archegonia, antheridia or both. In heterosporous plants, the spores are genetically different, carrying genes for either maleness or femaleness. Megaspores have genes that determine the female mating type and microspores the genes for the male mating type. These spores germinate and form gametophytes that will be either male or female and, since haploid, only the genes for one sex. When haploid eggs are produced by mitosis, they will contain a gene that determines "female"; when haploid sperm are produced by mitosis, they will contain a gene that determines "male". The fusion of a male sperm and a female egg (syngamy) will result in a zygote that contains both male and female genes and will grow to become a diploid sporophyte with both "sex genes". During the formation of spores by meiosis in the sporophyte, these sex genes will again be separated into haploid spores to produce the next generation of male and female gametophytes. If the female gene is represented by "a" and the male gene by "b", the megaspores will be represented by a and the microspores by b. Sperm can fertilize only eggs, so syngamy will always result in a zygote carrying both genes, a/b.
Protostele to Eustele Maybe the easiest to understand from structural considerations, is the advantage of more complex steles over the protostele. (Stele literally translates to upright pillar or column and refers to the vascular column of plants.) The earliest stele, the protostele, comprised only primary vascular tissue in a central core of the plant and could have a diameter no larger than the diameter of the apical meristem. This means that in the earliest land plants, such as Rhynia, the upright stems could be only a few milimeters in diamter. In some plants, like the Psaronius fern, the sporophyte first developed a massive apical meristem that allowed much larger protosteles. Siphonosteles are distinguished from protosteles by having a central pith surrounded by a cylinder of vascular tissue. This cylinder was more substantial and, like a soda straw, is structurally stronger than a simple stele. In addition, in many plants with siphonosteles, the vascular tissue of the cylinder retained the ability to divide, this becoming a vascular cambium, and the girth of the stem could enlarge. 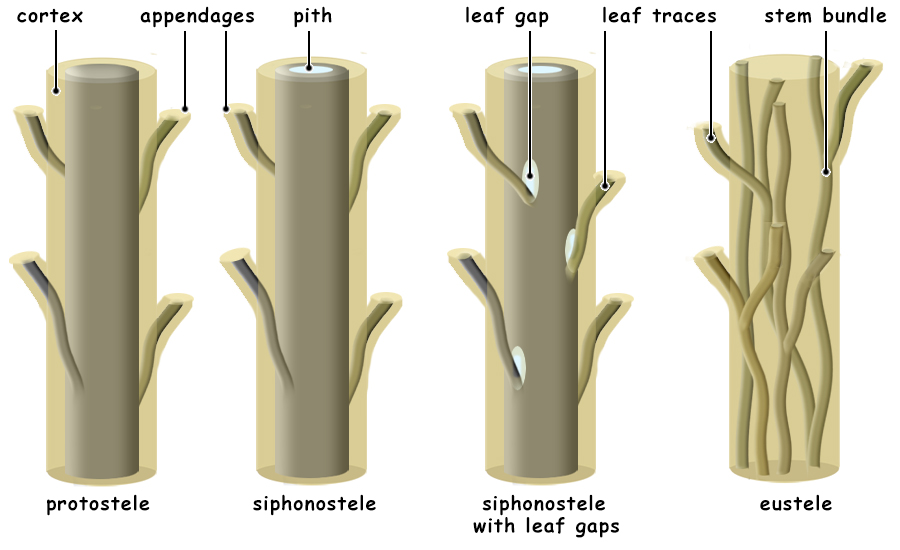 The most advanced eustele condition resulted in a stele that formed an anastomosing system of vascular bundles, each of which consisted of a bifacial vascular cambium. A bifacial vascular bundle produces new xylem (meta xylem) to the inside and new phloem (metaphloem) to the outside. This condition allows the trunk and stems to grow in girth and causes the older, outer tissues to crack as the interior enlarges. Along with this condition, parenchyma of the cortex gained the ability to divide and formed a layer of meristematic tissue outside the vascular bundles and and inside the outermost, dead layers of the bark. The vascular cambium and this cork cambium are the only tissues of the trunk and stem that can divide. Because the vascular cambium produces the newest cells to the exterior of the xylem and the interior of the phloem, the xylem continuously grows, layer upon layer, like dipping a candle. On the other hand, the newest phloem pushes the outer, older phloem outward, it cracks and the cork cambium "heals" the wounds created and forms the bark of these trees. There is no simple evolutionary line leading from plants with protosteles to plants with eusteles. Different evolutionary lineages adapted in many different ways to produce steles of various kinds. Enations to Leaves Like the steles of vascular plants, there is no singular evolutionary line leading from naked stems to megaphyllous leaves. Independent evolutionary steps occurred in the diverging plant phyla resulting in a diversity of leaf types. Also, like stelar evolution, this has resulted in great difficulties in developing a theory of how leaves and stems evolved . . . because they did so at different times and in different ways. It is best to consider the advantages conferred for each type of photosynthetic tissue type from a smooth photosynthetic epidermis, to small non-vascularized, spine-like enations, to the luxurient leaves of tropical plants. The first plants on land, the bryophytes, had phyllids that increased photosynthetic surfaces but that restricted them to aquatic habitats or wet habitats on land. Several adaptations of mosses allowed them to survive dessication, but never were able to achieve great heights. The problem is that large surface-to-volume (S/V) ratios favor evaporation and drying. To succeed on land, evaporation must be conquered. The earliest vascular land plants had simple, smooth stems which provided a favorable S/V ratio. Like the barrel cacti of deserts, a large volume and a small surface area allow them to "protect" an inner reservoir of water; any extended flat surface would lead to dehydration. Enations are small, non-vascularized scale-like outgrowths from the stems. Some hypothesize that their advantage was that it increased photosyhtetic surface are, while others propose that, like spines, they helped to protect the plant from insects which had invaded land long before plants. Both are probably true. 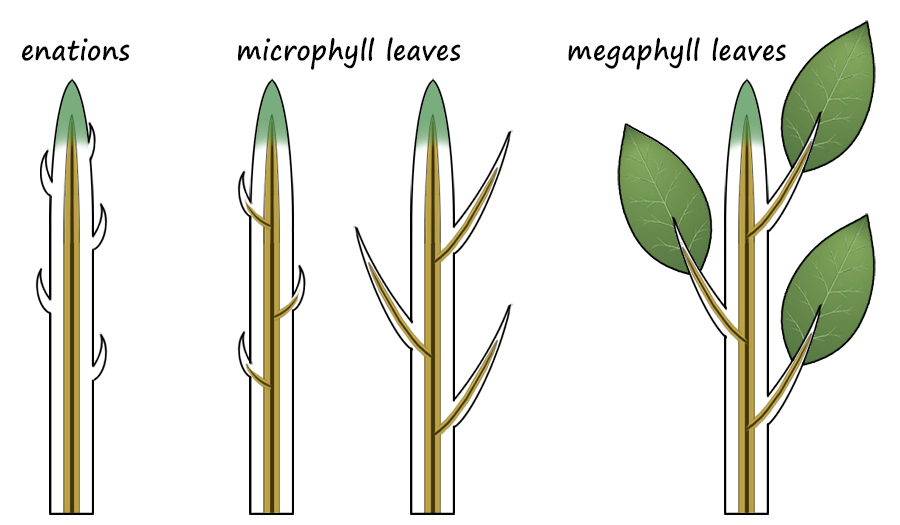 Once vascular tissue had evolved to better draw and distribute water from the soil, the luxury of having expanded photosynthetic surfaces was allowed and, in fact, leaves through evapotranspiration help to draw water from the soil. Leaves are defined as flatttened, photosynthetic structures attached to a stem and which contains an extension of vascular tissue from the stem. The simplest leaves have a single, unbranched vascular strand and are referred to as microphylls. And, although evolutionarily microphylls were indeed small, many plants developed enormous microphylls, some of which were more than three feet long, such as those of the lycophyte, Sigillaria. Megaphyll leaves are defined by leaves that have branching vascular bundles. Different theories exist for the transition from a leaf with a single vascular strand to one of many, anastomosing "veins" of vasular bundles. One such theory is the "telome theory" which states that leaves originated by the reduction in size of branches, the flattening of the plane of those branches, and the development of webbing between the reduced, flattened, branching system. This appears to be an untenable theory because it is far too complex when compared to the simple theory that enations became vascularized, expanded and the leaf vascular bundles branched . . . how simple is that? Vascular bundles branched from the stele to enter the leaf, why could they simply not continue to branch? Occam's Razor: of two theories that explain the same phenomenon, the simpler is more likely correct. |
| |